สารสื่อประสาท เซโรโทนิน Serotonin
Serotonin
โครงสร้างของ serotonin ประกอบด้วยหมู่ hydroxyl ซึ่งอยู่บนวงแหวนที่ 5 ของ indole nucleus และมี primary amine nitrogen เป็นตัวรับ H+ ทำให้ serotonin เป็นโมเลกุลที่มีขั้ว จึงไม่สามารถผ่านเยื่อหุ้มเซลล์และ blood brain barrier ได้ ดังนั้นการค้นพบ serotonin ในสมองจึงสามารถบ่งชี้ได้ว่า serotonin สามารถสร้างขึ้นในสมองและมีผลต่อหน้าที่สำคัญต่างๆ โดยเฉพาะผลต่อพฤติกรรมของสัตว์
ตำแหน่งของกลุ่มเซลล์ที่ทำหน้าที่สังเคราะห์ serotonin (serotonergic cell) ภายในสมอง
กลุ่มเซลล์ประสาทที่สร้าง serotonin จะอยู่ที่บริเวณแนวกลางของก้านสมองโดยมี axon กระจายอยู่ทั่วระบบประสาทส่วนกลาง ในปี ค.ศ. 1964 Dahlstrom และ Fuxe ได้พบกลุ่ม serotonergic cell เป็นจำนวนมากในบริเวณเซลล์ประสาท ที่เรียกว่า raphe nuclei ซึ่งสามารถสร้าง serotonin ออกมาได้ กลุ่มเซลล์ประสาทนี้ สามารถจำแนกออกเป็น 9 กลุ่ม ได้แก่ B1 ถึง B9 (ตารางที่ 1) และ พบว่าไม่ใช่ทุกเซลล์ใน raphe nuclei ที่สามารถสร้าง serotonin ได้ นอกจากนี้ยังพบกลุ่มเซลล์ประสาทนอก raphe nuclei ที่สามารถสร้าง serotoninได้ด้วย

กลุ่มเซลล์ประสาทที่สร้าง serotonin ที่ตำแหน่งต่าง ๆ ภายในสมอง
B1 --->
Raphe pallidus nucleus, caudal ventrolateral medulla
B2 --->
Raphe obscurus nucleus
B3 --->
Raphe magnus nucleus, rostral ventrolateral medulla, lateral paragigantocellular reticular nucleus
B4 --->
Raphe obscurus nucleus, dorsolateral part
B5 --->
Median raphe nucleus, caudal part
B6 --->
Dorsal raphe nucleus, caudal part
B7 --->
Dorsal raphe nucleus principal, rostral part
B8 --->
Median raphe nucleus, rostral main part; caudal linear nucleus; nucleus pontis oralis
B9 --->
Nucleus pontis oralis, supralemniscal region
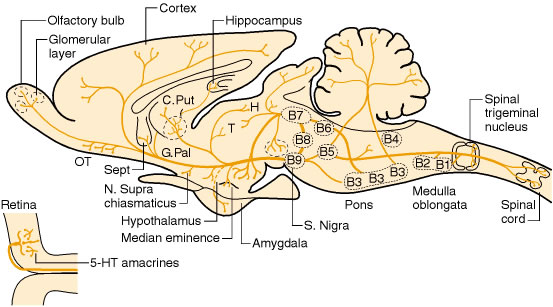
แสดงตำแหน่งและแขนงหลักของเซลล์ประสาทที่สร้าง serotonin ภายในระบบประสาทส่วนกลาง (OT = olfactory tuberculum; Sept = septum; C. Put = nucleus caudate - putamen; G. Pal = globus pallidus; T= thalamus; H = habenula; S. Nigra = substantia nigra) (ที่มา : Siegel, J. G. et al., 1999)
จากการศึกษาทางกายวิภาคพบว่า dorsal raphe, median raphe และ B9 cell มีแขนงประสาทขาขึ้นไปยังบริเวณสมองส่วนหน้าซึ่งเริ่มจาก midbrain ทั้งนี้ raphe nuclei จะให้เส้นทางหลัก ๆ 2 เส้นทาง คือ dorsal paraventricular path และ ventral tegmental radiation ทั้ง 2 เส้นทางนี้จะไปรวมกันที่ caudal hypothalamus ที่มี axon ของเซลล์ประสาทที่ผลิต dopamine และ noradrenaline ผ่านเข้ามายังบริเวณนี้
กลุ่มเซลล์ประสาทที่สร้าง serotonin กลุ่มอื่นๆได้แก่ B1-B4 จะอยู่ในส่วน caudal midpons จนถึงส่วน caudal medulla กลุ่มนี้จะให้แขนงไปยังส่วนของก้านสมองและไขสันหลัง โดยเฉพาะในส่วนของไขสันหลังจะได้รับแขนงประสาท เป็นจำนวนมาก โดยแขนงประสาทขาลงของเซลล์ประสาทที่สร้าง serotonin จะมี 3 เส้นทางหลัก ๆ ได้แก่
1. B3 (raphe magnus nucleus) ไปยัง laminae I,II ของ dorsal horn ของไขสันหลัง
2. B2 (raphe obscurus nucleus) ไปยัง lamina IX ของ ventral horn
3. B3 (rostral ventrolateral medulla และ lateral paragigantocellular reticular nucleus) ไปยัง interomediolateral cell
ส่วนแขนงประสาทจากกลุ่มเซลล์ B1 (caudal ventrolateral medulla) นั้นยังไม่มีข้อมูลแน่ชัด
การสังเคราะห์ serotonin
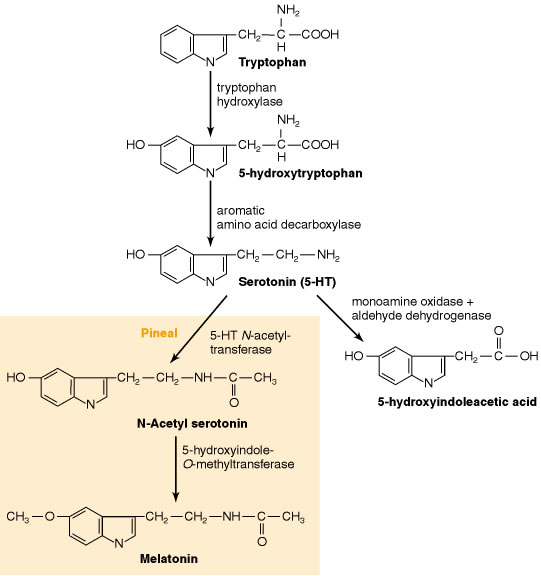
เซลล์บางชนิดอาจมี serotonin อยู่แต่ไม่สามารถสังเคราะห์ serotonin ได้ เช่น เกล็ดเลือดไม่สามารถสร้าง serotonin ได้เองแต่มันสามารถสะสม serotonin จากพลาสมาโดยกระบวนการ active transport ส่วนกลุ่มเซลล์ที่สามารถสร้าง serotonin ได้ คือ กลุ่มเซลล์สมองซึ่งมี การสังเคราะห์แบ่งเป็น 3 ขั้นตอน (ภาพที่ 7) ดังนี้
ขั้นที่ 1 นำกรดอะมิโน L-tryptophan จากกระแสเลือดเข้าสู่เซลล์สมองโดยวิธี facilitated diffusion ซึ่งแหล่งของ tryptophan ก็คือ อาหารโปรตีนต่างๆ tryptophan จะถูกขนส่งไปยังสมองโดยอาศัยตัวนำ (carrier) ชนิดเดียวกับ neutral amino acid อื่นๆ ได้แก่ phenylalanine, leucine และ methionine ดังนั้นจึงทำให้ tryptophan ต้องแข่งขันกับ neutral amino acid ตัวอื่นๆเพื่อเข้าจับกับตัวนำ ส่งผลทำให้ปริมาณของ serotonin ที่ถูกสังเคราะห์ลดลงได้ถ้าหากมีปริมาณของ tryptophan น้อย และส่งผลทำให้พฤติกรรมบางอย่างเปลี่ยนไป
ขั้นที่ 2 จัดเป็นขั้นกำหนดอัตราการเกิดปฏิกิริยา (rate-limiting step) โดยเซลล์ที่สร้าง serotonin จะมีเอนไซม์ L-tryptophan-5-monooxygenase หรือ tryptophan hydroxylase ทำหน้าที่เปลี่ยน tryptophan เป็น
5-hydroxytryptophan (5-HTP) โดยเอนไซม์นี้ต้องการออกซิเจน (O2) และ pteridine cofactor คือ L-erythro-tetrahydrobiopterin (BH4) เพื่อใช้ในการกระตุ้นเอนไซม์ให้สามารถทำงานได้ ออกซิเจน 1 อะตอมจะถูกใช้ในการสร้าง 5-HTP และอีก 1 อะตอมจะถูกรีดิวซ์(reduce) เป็นน้ำ pteridine cofactor จะเป็นตัวให้อิเลคตรอน ทำให้ได้สารกึ่งกลางที่ไม่เสถียร คือ quinonoid dihydrobiopterin แล้วเปลี่ยนเป็น quinonoid tetrahydrobiopterin (quinonoid BH4) ดังเช่นสมการข้างล่าง
L-tryptophan + BH4 + O2 ====>> L -5-HTP + quinonoid BH4 + H2O
ขั้นที่ 3 5-HTP จะถูกเปลี่ยนเป็น serotonin (5-HT) โดยเอนไซม์ aromatic L-amino acid decarboxylase (AADC) เอนไซม์นี้ไม่ได้พบเฉพาะในเซลล์ที่สร้าง serotonin เท่านั้นแต่ยังพบในเซลล์ที่สร้าง catecholamine ด้วย ซึ่งเอนไซม์จะทำหน้าที่เปลี่ยน 3,4-dihydroxyphenylalanine (dopa) เป็น dopamine แต่การเลือกใช้ 5-HTP หรือ dopa เป็นสารตั้งต้นของ AADC นั้น ขึ้นอยู่กับปริมาณความเข้มข้นของสารตั้งต้น cofactor และระดับ pH ดังนั้นถ้าในภาวะที่มี 5-HTP ในปริมาณต่ำ จะมีผลต่อปริมาณ serotonin ภายในสมอง และอีกสาเหตุหนึ่งที่ทำให้ปริมาณ serotonin ในสมองไม่อยู่ในระดับปกติ อาจเกิดจากมีเอนไซม์ไปจับบริเวณเซลล์ที่สร้าง catecholamine มากกว่าปกติ
การเก็บสะสมและการหลั่ง serotonin
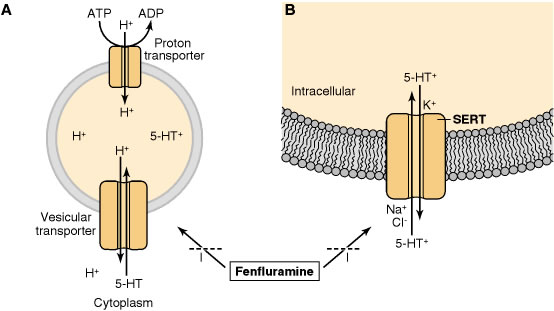
การสะสม serotonin ใน vesicle อาศัยขบวนการ active transport จาก cytoplasm โปรตีนที่ทำหน้าที่ขนส่งอาศัย ความแตกต่าง ทางศักย์ไฟฟ้าและเคมี (electrochemical gradient) และกระบวนการของเอนไซม์ H+-ATPase เป็นตัวผลักดันให้เกิด การขนส่ง โดยเป็นการแลกเปลี่ยน serotonin กับ H+ จึงทำให้ serotonin ถูกส่งเข้าสู่ vesicle และ H+ ก็จะถูกปล่อยออกสู่ cytoplasm
การเก็บสะสม serotonin ภายใน vesicle ไม่ต้องอาศัย ATP แต่จะมีโปรตีนที่มีความจำเพาะต่อ สารสื่อประสาท ตัวนี้อยู่ภายในเซลล์ที่ทำ หน้าที่สร้าง serotonin เรียกว่า serotonin binding protein (SBP) โดยโปรตีนนี้จะใช้ Fe2+ ช่วยในการจับกับ serotonin
การหลั่ง serotonin เกิดขึ้นโดยกระบวนการ exocytosis โดยใช้ serotonin transporter (SERT) เป็นตัวช่วยในการขนส่งเนื่องจาก serotonin เป็นโมเลกุลที่มีขั้ว จึงทำให้มันไม่สามารถผ่านเยื่อหุ้มเซลล์ โดยวิธี simple diffusion ได้ serotonin จึงถูกเก็บไว้ใน vesicle ซึ่งภายในจะมี serotonin-binding protein (SBP) และจะถูกหลั่งออกมาพร้อมกันกับ serotonin ด้วย
การสลาย serotonin
เอนไซม์ monoamine oxidase (MAO) จะเปลี่ยน serotonin ให้เป็น 5-hydroxy indoleacetaldehyde ซึ่งจะถูก oxidize ต่อโดยเอนไซม์ aldehyde dehydrogenase และ NAD+ ได้เป็น 5-hydroxyindoleacetic acid (5-HIAA) ซึ่งเป็นรูป metabolite ของ serotonin
เอนไซม์ MAO มี 2 isoform ได้แก่ MAO-A และ MAO-B โดยตัวยับยั้งของเอนไซม์ MAO-A คือ clorgyline หรือ moclobemido และตัวยับยั้งของเอนไซม์ MAO-B คือ deprenyl
ตัวรับสำหรับ serotonin ภายในสมอง
1. 5-HT 1A receptor
สามารถพบตัวรับนี้ได้มากที่สมองส่วน hippocampus, septum, amygdala, hypothalamus และ neocortex จากข้างต้น พบว่า ตำแหน่งที่ตัวรับเหล่านี้อยู่ส่วนใหญ่เป็นบริเวณของระบบลิมบิก (limbic system) ที่เกี่ยวข้องกับการควบคุมอารมณ์ นอกจากนี้ยังพบว่ าการทำลายเซลล์ประสาทที่เกี่ยวข้องกับ serotonin โดย neurotoxin 5,7-dihydroxytriptamine (5,7-DHT) ไม่สามารถไปลดจำนวนของ 5-HT 1A receptor ในสมองส่วนหน้า (fore brain) ลงได้ แสดงให้เห็นว่าตัวรับ 5-HT 1A นั้นอยู่บริเวณ postsynapse
การพบ receptor ชนิดนี้ในส่วนของ neocortex ทำให้คาดว่าน่าจะเกี่ยวข้องกับ ความสามารถในการรับรู้และการรวบรวมข้อมูลของสมอง ส่วน cortex 5-HT 1A receptor ยังสามารถพบได้ในกลุ่มเซลล์ที่ทำหน้าที่สร้าง serotonin เช่น dorsal และ medial raphe nuclei ซึ่ง 5-HT 1A receptor จะทำหน้าที่เป็นตัวรับของ serotonin ที่หลั่งออกมาจากปลายประสาทของกลุ่มเซลล์ที่ทำหน้าที่สร้าง serotonin การกระตุ้นที่ตัวรับบริเวณนี้จะไปลดการทำงานของเซลล์ประสาทที่เกี่ยวข้องกับการหลั่ง serotonin และ ลดการปล่อย serotonin จากปลายประสาทของเซลล์ประสาทเหล่านั้น
2. 5-HT 1B และ 5-HT1D receptor
ตัวรับ 5-HT 1B พบได้ในสมองของหนู rat และสมองของหนู mice ส่วน 5-HT 1D receptors พบได้ในสมองของโค และ สมองของมนุษย์ โดยจะพบจำนวนมากใน basal ganglia บางส่วนใน globus pallidus และ substantia nigra
ตัวรับ 5-HT 1B และ 5-HT 1D จะอยู่ที่บริเวณ presynapse ของเซลล์ประสาทที่เกี่ยวกับการสร้างและปล่อย serotonin โดยตัวรับทั้งสองจะทำหน้าที่ควบคุมการปล่อย serotonin ออกจากเซลล์ประสาทนี้ การปล่อยของ serotonin จาก dorsal raphe nucleus ก็อยู่ภายใต้การควบคุม ของ receptor สองตัวนี้เช่นกัน นอกจากนี้ยังพบตัวรับทั้งสองนี้ในส่วน postsynapse อีกด้วย
เป็นไปได้ว่าตัวรับทั้งสองนี้อาจมีส่วนในการควบคุมการปล่อยของสารสื่อประสาทตัวอื่น เช่น acetylcholine ใน hippocampus และ dopamine ใน prefrontal cortex
3. 5-HT 2A receptor
พบได้ในบริเวณผิวนอกทั่วไปของสมอง และ พบมากในส่วนcortex ด้านหน้าบริเวณ claustrum ซึ่งเป็นบริเวณที่ติดต่อกับส่วน visual cortex, limbic system, basal ganglia และ olfactory nuclei และยังพบตัวรับนี้ที่บริเวณ postsynapse ของเซลล์ประสาทชั้นในของสมองส่วน cerebral cortex (intrinsic cortical neuron)
4. 5-HT 2C receptor
พบมากที่ epithelial cell ของ choroid plexus และยังพบทั่วไปในสมอง โดยเฉพาะบริเวณ limbic system, hypothalamus, hippocampus, septum, neocortex และ บริเวณที่เกี่ยวข้องกับการสั่งงานทางพฤติกรรม คือ substantia nigra และ globus pallidus
เมื่อตัวรับ 5-HT 2C จับกับ serotonin สามารถควบคุมส่วนประกอบและปริมาณของcerebrospinal fluidได้ แต่จากการทดลองเรายังไม่พบสารเสริมการทำงาน (agonist) และ สารยับยั้งการทำงาน (antagonist) ที่แท้จริงของตัวรับนี้ จึงทำให้ยังไม่ถึงทราบหน้าที่อื่นๆอย่างแน่ชัด
5. 5-HT 3 receptor
บริเวณที่พบตัวรับนี้มากสุดในสมอง คือ area postrema และมักพบในเซลล์ประสาทรอบนอก (peripheral neuron) เช่น superior cervical ganglion , vagus nerve , substantia gelatinosa ของ spinal cord serotoninสามารถควบคุมกลไกการตอบสนองต่อความเจ็บปวดได้โดยผ่านทาง 5-HT 3 receptor และ ยังกระตุ้นการปล่อยของ substance P ใน spinal cord ด้วย ส่วนผลต่อระบบลิมบิกนั้นมีผู้ศึกษาโดยให้สารยับยั้งการทำงานของ 5-HT3 receptor (5-HT3 receptor antagonist) ผลการทดลองพบว่าสามารถลดความวิตกกังวล และมีผลกระทบต่อการรับรู้ด้านความเจ็บปวดด้วย
ตัวรับ 5-HT3 เป็นตัวช่วยให้เกิด depolarization ของเซลล์ประสาทที่ทำหน้าที่ส่งผ่าน serotoninได้ นอกจากนี้ตัวรับ 5-HT3 ยังสามารถควบคุมเซลล์ประสาทที่ผลิต dopamine และ acetylcholine ด้วย
ในสมองส่วน ventral tegmentam, cortex และ ที่สมองส่วน hippocampus พบว่ามีเซลล์ประสาทที่มีตัวรับ 5-HT3 อยู่ คือ เซลล์ประสาทกลุ่ม GABAnergic
6. 5-HT 4 receptor
พบมากใน striatum , substantia nigra, olfactory tubercle และ hippocampus ตัวรับ 5-HT 4 เป็นตัวช่วยเพิ่มประสิทธิภาพในการหลั่ง dopamine ที่ส่วน striatum ถึงแม้ว่าจะไม่พบตัวรับ 5-HT 4 ในบริเวณส่วนปลายของเซลล์ประสาทที่มีการหลั่ง striatal dopamine นี้ก็ตาม
7. 5-HT 7 receptor
การศึกษาในหนูพบตัวรับชนิดนี้ ภายในสมองส่วน cortex, septum, thalamus, hypothalamus, amygdala และ superior colliculus ส่วนในอวัยวะอื่นๆ ตัวรับนี้จะทำหน้าที่เป็นสื่อให้เกิดการคลายตัวของกล้ามเนื้อเรียบภายในหลอดเลือด